Importancia de las leguminosas en la alimentación de rumiantes

Dervin Dean Engormix
INTRODUCCION
En el trópico las gramíneas ofrecen la fuente más barata de nutrientes disponibles para la alimentación animal, pero existen factores limitantes muy importantes, debido al bajo aporte proteico, con altos valores de fibra, baja digestibilidad y frecuentemente con graves deficiencias minerales, y consecuentemente, son comunes los bajos niveles de producción, ya que la productividad de los animales a pastoreo está básicamente determinada por el consumo diario de energía y proteína (Ruiz &Vásquez, 1983).
El suministro de fuentes proteicas es relativamente costoso; en tal sentido, se ha venido trabajando en la búsqueda de alternativas que resulten económicas y viables.
Las leguminosas son especies capaces de sintetizar altos niveles de proteína cruda (PC), con una tasa relativamente baja de disminución de este componente en la medida que la planta madura (Rincón, 2011), comparada con especies de gramíneas tropicales.
En el Cuadro 1 (Clavero, 2011) se presentan valores nutricionales de diferentes especies de leguminosas, donde se observa que no solo el contenido proteico es elevado, sino también los valores de digestibilidad reportados superan el de muchas gramíneas tropicales, por lo que el aporte energético de las leguminosas pudiera ser mayor que el de las gramíneas.
Según Clavero (2011), además de representar buenas fuentes de proteína, estos materiales también son una importante fuente de minerales, tales como calcio con contenidos que se encuentran en un rango de 0,24-1,90%, mientras para fosforo el rango es 0,19-0,40%, magnesio 0,10-0,47%, sodio 0,05-0,14%, potasio 0,32-2,75%, cobre 17-27 ppm, zinc 28-43 ppm y manganeso 31-117 ppm.
De acuerdo a McDowell et al. (1989), la mayoría de los arboles forrajeros contienen suficientes macro y micro minerales para cubrir los requerimientos de la ganadería en las regiones tropicales.
Todas estas características las convierten en recursos alimenticios de alto potencial en la ganadería de carne y leche tropical, como fuente de proteína de bajo costo, y cuyo uso como fuentes forrajeras ayudarían a incrementar la calidad de la dieta de los animales.
Cuadro1. Valor nutricional de algunas especies leguminosas
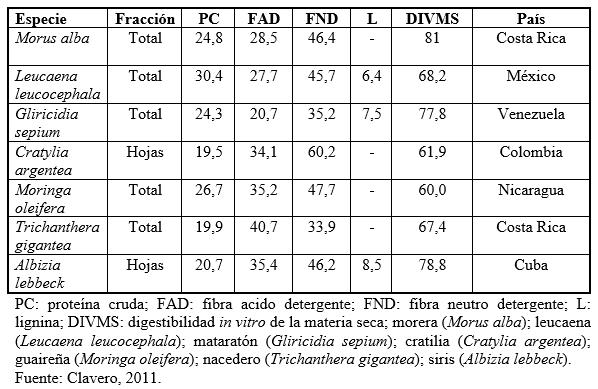
El objetivo del presente capitulo es de resaltar las potencialidades y ventajas que podrían representar las leguminosas tropicales, como fuentes proteicas, en los sistemas de producción de rumiantes en el trópico.
EFECTO DEL CONSUMO DE LEGUMINOSAS SOBRE LA FERMENTACIÓN RUMINAL
Diversos estudios han demostrado que el consumo de especies leguminosas mejora la degradabilidad de la fibra, el consumo de materia seca y aumenta la población de microorganismos ruminales, los cuales son indispensables para que los rumiantes mejoren el aprovechamiento de los recursos fibrosos.
Chanthakhounet al. (2011) evaluaron el efecto del consumo de tres niveles crecientes (0, 300, 600 y 900g MS/d) de heno de chi xiao dou (Phaseolus calcaratus) (con 18% de PC) sobre la digestibilidad en búfalos de pantanos alimentados con residuos de cosechas de arroz y observaron que la suplementación con el heno de la leguminosa incremento linealmente el consumo total de materia seca (CMS), la digestibilidades aparentes de PC y materia orgánica (MO) y la producción de ácidos grasos volátiles (AGV) lo cual es una consecuencia del aumento en la tasa de fermentación de la dieta.
Resultados similares fueron obtenidos por Camero & Franco (2001) quienes utilizaron novillos Bos indicus que consumieron una dieta basal de heno de yaraguá (Hyparrhenia rufa) + pulitura de arroz y melaza, y fueron suplementados con henos de bucare ceibo (Erythrina poeppigiana) o mataratón (Gliricidia sepium) y un tercer grupo que recibió urea y observaron que la desaparición de materia seca a nivel ruminal aumento y el potencial de degradación fue mayor para los animales que consumieron las leguminosas comparados con los que consumieron urea.
Sin embargo, no detectaron diferencias entre los que consumieron alguna de las dos leguminosas.
También observaron un aumento de entre 15 a 24% en la concentración de AGV en el licor ruminal entre los que consumieron las leguminosas comparados con los que consumieron urea.
Por su parte Abreu et al. (2004), alimentaron ovinos West African adultos con heno de baja calidad de pasto llanero (Brachiaria dictyoneura) (3,7% PC) solo o mezclado en una relación 3:1 con cratilia (Cratylia argentea) (18,6% de PC) y observaron en los animales que recibieron en su dieta la mencionada leguminosa, que el consumo de MO y de PC se incrementaron en 21% y 130%, respectivamente.
Weisbjerg & Soegaard (2008) sugieren que el mayor consumo de leguminosas pudiera ser, entre otras causas, a diferencias en la estructura química y el contenido de fibra, comparadas con las gramíneas.
Según estos autores, las leguminosas tienen una menor concentración de fibra neutro detergente (FND) total, sin embargo a su vez poseen una mayor concentración de lignina que las gramíneas.
Pero debido a que esta lignina está ligada a una baja proporción de la FND indigerible, existe una mayor proporción de FND digerible en las leguminosas que en las gramíneas, seria la razón para que las primeras tengan mayor digestibilidad total de la MO que las segundas.
En las leguminosas, la lignina se encuentra solo en el xilema, por lo que este es completamente indigerible, mientras que en el resto de los tejidos no hay lignina y las paredes celulares son completamente digeribles (Wilson & Kennedy, 1996).
Sin embargo, en las gramíneas la lignina se encuentra distribuida en todos los tejidos de la planta, excepto en el floema, por lo tanto, la menor cantidad de lignina que poseen las gramíneas protegen una mayor cantidad de paredes celulares de la degradación ruminal, lo que provoca que la tasa de digestión de la pared celular sea menor que en las leguminosas (Buxton & Russell, 1988).
Evaluaciones realizadas sobre cinética de fermentación ruminal, han mostrado que las diferencias en la estructura y contenido de las paredes celulares mencionadas anteriormente, provocan un aumento en la tasa de pasaje de las leguminosas, explicando la mayor capacidad de consumo de estas últimas, comparadas con las gramíneas (Kuoppala et al., 2009).
Allen (1996) estima que al incrementarse la densidad de las partículas en el rumen, se acorta el tiempo de retención, y el aumento de la gravedad del tamaño de partícula es probablemente mayor en leguminosas que en gramíneas, debido a la tasa de digestión más acelerada de la FND potencialmente digestible, lo cual promueve un aumento en la tasa de pasaje.
Un estudio reveló que la concentración de nitrógeno amoniacal (N-NH3) en el licor ruminal de ovinos, alimentados con pasto llanero (3,7% PC), se incrementó de 2,40 a 8,43 mg/100 ml cuando el pasto fue mezclado en una relación 3:1 con cratilia (18,6% de PC, Abreu et al., 2004).
La concentración de N-NH3 es de suma importancia para rumiantes alimentados con dietas ricas en fibra, ya que las bacterias fibrolíticas usan este sustrato nitrogenadode preferencia para la síntesis de su proteína; Boucher et al. (2007) determinaron que estas crecían a un mayor tasa cuando la concentración de N-NH3 era de alrededor de 12,8 mg/100 ml.
En el trabajo de Abreu et al. (2004) también se observó mayor cantidad de nitrógeno total (+56%) y nitrógeno microbial (+28%) a nivel del duodeno de los animales que consumieron la leguminosa.
Chanthakhoun et al.(2011) también observaron un incremento lineal en la retención de N al igual que en la eficiencia de la síntesis de proteína microbial con niveles creciente de heno de leguminosa en búfalos. Estos mismos investigadores demostraron que el consumo de heno de leguminosa puede incrementar las poblaciones de bacterias, protozoos y de hongos en búfalos, mientras que la producción de gas metano en el rumen disminuyó linealmente en la medida que aumentaba la proporción de la leguminosa en la dieta.
Según Moore et al. (1999) probablemente el elemento nutricional que más afecta el consumo voluntario (CV) de forrajes es la PC, y cuando la concentración de este nutriente en la ración es inferior a 8%, el CV tiende a deprimirse, ya que bajo estas condiciones el crecimiento de los microorganismos ruminales pudiera estar limitado por la cantidad de nitrógeno disponible.
Esto probablemente sea la causa principal por lo que las leguminosas forrajeras pudieran mejorar el patrón de fermentación ruminal, ya que pueden proveer el nitrógeno necesario para un crecimiento microbiano adecuado, con lo cual se incrementaría el aprovechamiento de los recursos fibrosos que consume el rumiante.
EFECTO DEL CONSUMO DE ESPECIES LEGUMINOSAS EN LA RESPUESTA ANIMAL
Diferentes estudios han mostrado los efectos positivos de la suplementación con especies leguminosas sobre la producción de leche en diferentes especies rumiantes.
Razz & Clavero (1997) evaluaron el efecto de remplazar 2 kg/vaca/d de alimento balanceado por 2 kg/vaca/d de mataratón en vacas lecheras consumiendo pasto barrera (Brachiaria decumbens) y observaron que la producción de leche se incremento en 12,1 y 13,9% en las vacas que consumieron heno de la leguminosa vs las que consumían los 2 kg de alimento y las que consumían solamente gramínea, respectivamente.
Steinshamn (2010) por su parte, realizó una extensa revisión de evaluaciones que se han llevado a cabo para evaluar el efecto de la suplementación con leguminosas sobre producción de leche y al compararlo con dietas basadas en gramíneas observo que el rango en aumento en producción de leche fue de entre 1,1 y 1,6 kg/d para las diferentes leguminosas.
También reporta aumentos en el consumo de materia seca en los diferentes ensayos.
Diferentes estudios también han evaluado el efecto de leguminosas con potencial forrajero durante la fase de crecimiento en diferentes especies de rumiantes y se han obtenido resultados positivos.
Combellas et al.(1999) evaluaron en dos experimentos, el efecto de leucaena y mataratón sobre la ganancia en peso de ovinos de la raza West African en crecimiento, recibiendo como dieta basal pasto de corte a voluntad.
En el primer experimento utilizaron dos lotes, uno fue suplementado con 130 g/animal/d de harina de ajonjolí (HA) y el otro con leucaena fresca (LF).
Se obtuvieron ganancias en peso de 53 g/d para HA y 87 g/día para LF, siendo mayores las ganancias de las corderas suplementadas con follaje de leucaena.
En el segundo ensayo se utilizaron 3 lotes, donde los animales fueron suplementados durante 42 días con 130 g/d de harina de pescado (HP, lote 1), mataratón (M, lote 2) y 130 g/día de harina de pescado + mataratón (HPM, lote 3).
Las ganancias en peso fueron respectivamente para HP, M y HPM de 83, 66 y 98 g/d, siendo inferiores las de las corderas suplementadas solo con M a las de las suplementadas con HPM.
Los investigadores concluyen que el uso del follaje de ambas leguminosas es factible como suplemento proteico para ovinos en crecimiento.
Por su parte Verdolak & Zorate (2008) evaluaron 5 tratamientos, agregando 25% de salvado de trigo, ó 25% de alfalfa (Medicago sativa), ó 25% de leucaena, ó 25% de guajillo (Desmanthus virgatus) ó 25% de zapatico de la reina (Clitoria ternatea), a dietas para corderas mestizas Dorper en crecimiento, basadas en heno de pasto buffel (Cenchrus ciliaris), y no observaron diferencias significativas en la conversión alimenticia entre los 5 tratamientos.
Los valores encontrados para la conversión alimenticia en dicha evaluación fueron de 9,7; 11,1; 10,4; 10,0 y 11,9 kg MS por kg de aumento obtenido con el uso de raciones con salvado, alfalfa, leucaena, guajillo y zapatico de la reina, respectivamente.
Benavides-Calvache et al. (2010), evaluaron durante siete meses en terneros mestizos F1 (Brahmán x Holstein, 50%) x razas taurinas, el efecto de dos tratamientos: 1. pastoreo en potreros de pasto mulato (Brachiaria hibrido) y suplementados con una mezcla de gallinaza, salvado de maíz y caña de azúcar, y 2. pastoreo en potreros de pasto mulato en asociación con cratilia (49% de mulatoy 51% de cratilia).
Se encontraron ganancias de peso de 59,1 vs 68,7 kg/animal para los grupos 1 y 2 respectivamente, durante el periodo experimental.
Los resultados presentados previamente, son evidencia que el uso de leguminosas en las dietas para rumiantes, sirve para corregir algunas deficiencias nutricionales, especialmente proteicas, que son comunes en las gramíneas tropicales.
La conveniencia de utilizar leguminosas en la fase de crecimiento se deriva de la necesidad de usar dietas altas en proteínas, ya que la síntesis de tejido magro (musculo y órganos internos) es elevada durante esa fase y la PC cruda aportada por las leguminosas permitiría compensar los bajos niveles proteicos de las gramíneas.
PRESENCIA DE FACTORES ANTINUTRICIONALES EN LEGUMINOSAS
Un factor antinutricional (FAN) puede definirse como aquella sustancia sintetizada en los alimentos de forma natural,que ejerce efecto contrarios a una nutrición óptima, reduciendo el consumo e impidiendo la digestión, la absorción y la utilización de nutrientes por el animal (Kumar, 2013).
Son sustancias naturales no fibrosas generadas por el metabolismo secundario de las plantas, como un mecanismo de defensa ante el ataque de mohos, bacterias, insectos y pájaros.
Su naturaleza, mecanismos de acción y potencia de sus efectos son muy variados y tienen una amplia distribución en el reino vegetal.
En el Cuadro 2 se presenta una lista de los FAN que pudieran estar presentes en las leguminosas, tanto arbóreas como arbustivas, utilizadas como fuentes forrajeras en la alimentación de rumiantes.
En general, frente a condiciones severas (p.e. climas áridos), las plantas tienden a aumentar sus defensas, sobre todo de tipo cualitativo (alcaloides, glucósidos cianogenéticos, entre otros), ya que en estos casos, les resulta mucho más difícil regenerar los tejidos dañados por los herbívoros (Jhonson et al., 1985).
Esto representa un riesgo potencial para los animales, ya que aumentan las incidencias de intoxicación en el rebaño cuando se enfrenta a la escasez de otros recursos vegetales (James et al., 1992).
Cuadro 2. Efecto de los factores antinutricionales en rumiantes
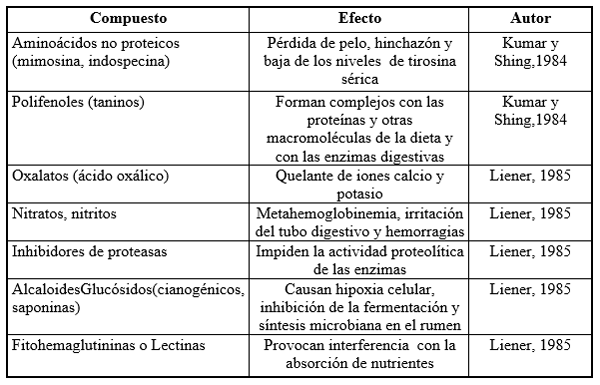
La fermentación ruminal puede modificar muchos factores antinutricionales a formas menos toxicas para el metabolismo de los mamíferos y por tanto reducir la susceptibilidad de los rumiantes a muchos de estos compuestos (Dixon & Hosking, 1992).
El medio ruminal representa un lugar eficiente de detoxificación para un amplio rango de compuestos secundarios de las plantas (terpenos, fenoles, ácidos, entre otros), de modo que la toxicidad de las plantas consumidas por los rumiantes puede ser modificada significativamente después de los cambios químicos sufridos por los compuestos potencialmente tóxicos en el rumen (Domínguez-Bello, 1996).
La proporción gramínea:leguminosa en dietas para rumiantes fue estudiada por Norton (1994) y este investigador determinó que al elevarse porcentaje de leucaena por encima de 40% en la ración, no se evidenciaron mejoras en el consumo voluntario ni en la digestibilidad de la ración, lo que podría indicar que una proporción 60:40 gramínea-leguminosa pudiera ser lo adecuado en dietas para especies rumiantes.
CONCLUSIÓNES
Las leguminosas representan una alternativa importante para suplir ciertas deficiencias nutricionales de rumiantes en condiciones tropicales, especialmente proteicas.
Estas ejercen su efecto positivo a partir del rumen, donde incrementan la capacidad de fermentación de la fibra y potencian el consumo voluntario.
El alto contenido de proteína cruda de la mayoría de las leguminosas promueve un aumento en la tasa de crecimiento y en producción láctea en diferentes especies rumiantes.
Sin embargo, la presencia de factores antinutricionales en la generalidad de estas especies pudiera limitar la inclusión en altas proporciones de estas especies en dietas para rumiantes.
REFERENCIAS BIBLIOGRAFICAS
Abreu A, Carulla JE, Lascano CE, Diaz TE, Kreuzer M, Hess HD. 2004. Effects of Sapindus saponaria fruits on ruminal fermentation and duodenal nitrogen flow of sheep fed a tropical grass diet with and without legume. J Anim Sci 82:1392-1400.
Allen MS. 1996. Physical constraints on voluntary intake of forages by ruminants. J Anim Sci 74: 3063-3075.
Benavides-Calvache CA, Valencia-Murillo M, Estrada-Álvarez J. 2010. Efecto de la veranera forrajera (Cratylia argentea) sobre la ganancia de peso de ganado doble propósito. Vet Zootec 4: 23-27.
Boucher SE, Ordway RS, Whitehouse NL, Lundy FP, Kononoff PJ, Schwab CG. 2007. Effect of incremental urea supplementation of a conventional corn silage-based diet on ruminal ammonia concentration and synthesis of microbial protein. J Dairy Sci 90: 5619-5633.
Camero, A. and M. Franco (2001). Improving rumen fermentation and milk production with legume tree fodder in the tropics. Agroforestry systems 51:157-166.
Chanthakhoun V, Wanapat M, Wachirapakorn C,Affiliations Wanapat S. 2011. Effect of legume (Phaseolus calcaratus) hay supplementation on rumen microorganisms, fermentation and nutrient digestibility in swamp buffalo. Livestock Sci 140: 17-23.
Clavero T. 2011. Agroforesteria en la alimentación de rumiantes en América Tropical. Revista de la Universidad del Zulia. Ciencias del Agro, Ingeniería y Tecnología 2: 11-35.
Combellas J, Ríos L, Osea A, Rojas J. 1999. Efecto de la suplementación con follaje de leguminosas sobre la ganancia en peso de corderas recibiendo una dieta basal de pasto de corte. Rev Fac Agron (LUZ) 16: 211-216.
Dixon RM, Hosking BJ. 1992. Nutritional value of grain legumes for ruminants. Nutr Res Rev5: 19-43.
Dominguez-Bello MG. 1996. Detoxification in the rumen. Ann Zootech 45: 323-327
Johnson AE, Molyneux RJ, Merrill GB. 1985. Chemistry of toxic range plants. Variation in pyrrolizidine alkaloid content of Senecio, Amsinckia, and Crotalaria species. J Agric Food Chem 33: 50- 55.
James LF, Nielsen DB, and Panter KE. 1992. Impact of poisonous plants on the livestock industry. J Range Manage 45: 3-8.
Kumar R, Shing M. 1984. Tannins: their adverse role in ruminant nutrition. J Agric Food Chem 32: 447-453.
Kumar R. 2013. Anti-nutritional factors, the potential risks of toxicity and methods to alleviate them. FAO, Agric and Cons Protect. Obtenida el 20 de agosto de 2013 de http://www.fao.org/docrep/003/t0632e/t0632e10.htm.
Kuoppala K, Ahvenjarvi S, Rinne M, Vanhatalo A. 2009. Effects of feeding grass or red clover silage cut at two maturity stages in dairy cows. 2. Dry matter intake and cell wall digestion kinetics. J Dairy Sci 92: 5634-5644.
Liener IE. 1985. Chemistry and Biochemistry of Legumes. Edit. Edward Arnold. India. pp 217-257.
McDowell LR, Conrad JH, Ellis GL, Loosli JK. 1989. Minerals for Grazing Ruminants in Tropical Regions. Academic Press, Inc. New York. USA. pp. 92.
Moore JE, Brant MH, Kunkle WE, Hopkins DI. 1999. Effects of supplementation on voluntary forage intake, diet digestibility, and animal performance. J Anim Sci 77 (2):122-135.
Norton BW. 1994. Tree legumes as dietary supplements for ruminants. In: Forage Tree Legumes in Tropical Agriculture. Gutteridge RC, Shelton HM (eds). CAB International. Wallingford, Oxford, UK. pp 192-201.
Razz R, Clavero T. 1997. Producción de leche en vacas suplementadas con harina de Gliricidia sepium. Arch Latinoam Prod Anim 5(1): 127-128.
Rincón J. 2011. Establecimiento y manejo de leguminosas arbóreas de importancia forrajera en zonas semiáridas de Venezuela. En: Innovación & Tecnología en la Ganadería Doble Propósito. González-Stagnaro C, Madrid-Bury N, Soto Belloso E (eds). Fundación GIRARZ. Ediciones Astro Data S.A. Maracaibo, Venezuela. Capítulo XXIX: 277-289.
Ruiz R, Vázquez CM. 1983. Consumo Voluntario de pastos y forrajes tropicales. En Ugarte J, Senra C (eds). Los Pastos en Cuba. Tomo 2. La Habana, Cuba. pp. 117-123.
Steinshamn H. 2010. Effect of forage legumes on feed intake, milk production and milk quality-a review. Anim Sci Papers and Reports 28: 195-206.
Verdoljak JO, Zórate P. 2008. Uso de Leguminosas Tropicales en la Alimentación de Ovinos de Pelo. INTA-Argentina. Informe de Investigación. Obtenido el 20 de agosto de 2013 dehttp://inta.gob.ar/personas/verdoljak.juan.
Weisbjerg MR, Soegaard K. 2008. Feeding value of legumes and grasses at different harvest times. In: Proceedings of 22nd General meeting of the European Grassland Federation. Hopkins A, Gustafson T, Nilsdotter-Linde N, Spörndly E (eds). Grassland Sci in Europe 13: 513-515.
Wilson JR, Kennedy PM. 1996. Plant and animal constraints to voluntary feed intake associated with fibre characteristics and particle breakdown and passage in ruminants. Aust J Agric Res 47: 199-225.
Publicado en “Manejo de Pastos y Forrajes Tropicales” correspondiente a la LXIII Reunión GIRARZ. Cuadernos Científicos GIRARZ 13 © 2013 Fundación GIRARZ
No hay comentarios:
Publicar un comentario